SNARE family members associate into stable complexes that direct membrane fusion events. The SNARE mediated process of vesicle Ginsenoside-F4 exocytosis is not only critical for phagocytosis but is also required for the release of inflammatory cytokines from macrophages and microglia. SNARE complexes prime vesicles for fusion with the plasma membrane and are composed of proteins from each of three separate families: a SNAP, a Syntaxin, and a VAMP. SNARE complexes are formed by cells in anticipation of membrane fusion events and upon stimulation, the complexes localize to the points of vesicle contact, facilitate membrane fusion, and then dissociate into their individual components that are recycled to form new complexes. Numerous studies in other labs have shown that a-syn may affect SNARE complex assembly either through direct interaction with members of the SNARE family, or by sequestering agents, such as arachadonic acid, which promote SNARE assembly and activity. We have recently developed a novel transgenic genomic mouse model in which wild type or the E46K mutant form of human asyn was expressed from a human bacterial artificial chromosome. This model is similar to that created by Kou et al. however, we focused on wild type and the E46K mutated form of a-syn rather than the A30P and A53T mutations described previously. The BAC construct contains the a-syn promoter and other potential regulatory regions including upstream, and 15 kb downstream sequences. Because a-syn expression is driven from its own promoter, not from a highly active exogenous promoter, this in vivo system may provide insight into human disease, because in humans duplication or triplication of the gene is sufficient to lead to early onset PD. In addition, 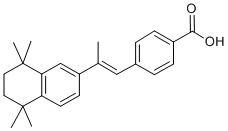 the potential impact of elevated asyn levels on multiple Sipeimine cellular and tissue systems can be assessed. In the work described here, we used these mice to examine the role of a-syn in innate immune cell function. We were particularly interested in macrophages and microglia because of their involvement in PD pathology and their intensive reliance on membrane dynamics for their central roles in cytokine release and phagocytic function. Our data demonstrates that increased expression of the a-syn protein has an inhibitory effect on the release of inflammatory cytokines, vesicular fusion, and phagocytic processes. Defective phagocytosis occurred in brain microglia and peripheral macrophages, either isolated and assayed in culture, or assayed in situ. Fibroblasts and monocytes isolated from sporadic PD patients and a patient carrying a triplication of the a-syn gene exhibited elevated a-syn levels, which correlated with defective phagocytic function. We also expand upon the molecular details regarding asyn interaction with membrane trafficking components, and demonstrate active recruitment of a-syn to a site of stimulated membrane fusion during phagocytosis. Our data further defines the role that a-syn plays in membrane vesicle dynamics and extends these findings into systemic processes beyond neuronal function, which may provide a valuable peripheral biomarker of elevated a-syn levels and dysfunctional vesicle dynamics. Our data suggest that there may be a previously unappreciated involvement of innate immune cell function in synucleinopathies, and that cells of the innate immune system provide an excellent, tightly regulated cellular model in which to study the effects of increased a-syn levels on vesicle function. Our data demonstrate that a-syn is expressed in innate immune cells and that the level of a-syn expression can regulate immune cell function.
the potential impact of elevated asyn levels on multiple Sipeimine cellular and tissue systems can be assessed. In the work described here, we used these mice to examine the role of a-syn in innate immune cell function. We were particularly interested in macrophages and microglia because of their involvement in PD pathology and their intensive reliance on membrane dynamics for their central roles in cytokine release and phagocytic function. Our data demonstrates that increased expression of the a-syn protein has an inhibitory effect on the release of inflammatory cytokines, vesicular fusion, and phagocytic processes. Defective phagocytosis occurred in brain microglia and peripheral macrophages, either isolated and assayed in culture, or assayed in situ. Fibroblasts and monocytes isolated from sporadic PD patients and a patient carrying a triplication of the a-syn gene exhibited elevated a-syn levels, which correlated with defective phagocytic function. We also expand upon the molecular details regarding asyn interaction with membrane trafficking components, and demonstrate active recruitment of a-syn to a site of stimulated membrane fusion during phagocytosis. Our data further defines the role that a-syn plays in membrane vesicle dynamics and extends these findings into systemic processes beyond neuronal function, which may provide a valuable peripheral biomarker of elevated a-syn levels and dysfunctional vesicle dynamics. Our data suggest that there may be a previously unappreciated involvement of innate immune cell function in synucleinopathies, and that cells of the innate immune system provide an excellent, tightly regulated cellular model in which to study the effects of increased a-syn levels on vesicle function. Our data demonstrate that a-syn is expressed in innate immune cells and that the level of a-syn expression can regulate immune cell function.