Plant volatiles can modulate both the degree of attraction and the physiological response to sex pheromone in males of many moth species. The olfactory background against which it is sensed can thus affect the behavioral response towards an odor. In the present study we asked how leaf and flower blends interact in nectar foraging female M. sexta hawk moths. Females visit plants for both nectar feeding and oviposition. Odor-based localization of flowers and potential host plants has been well described. Plant-derived leaf odors are typically discussed in the context of host-plant localization and oviposition site choice, whereas flower odors are studied from a foraging perspective. It has been shown that a few key compounds of the D. wrightii flower odor blend are necessary and sufficient to provoke directed upwind flights in Manduca. In the present study, we also show that a mixture of two components of the N. AbMole D-Pantothenic acid sodium attenuata flower bouquet provokes plume following and feeding behavior in a similar way. Flower blends are highly attractive stimuli for a hungry moth. Beyond attractiveness our data also show that the mixtures used convey odor identity to the foraging females since they were discriminated in a choice test using similar odor intensities. Next we asked how leaf background odors affect the attractiveness of flower odors. In order to focus rather on foraging than on oviposition behavior, we used unmated and hungry Manduca females. It has been described that aged unmated Manduca moths sometimes 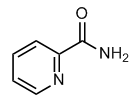 show oviposition behavior and lay unfertilized eggs. However, in agreement with the data reported by we never observed abdomen curling, which would indicate egg-laying behavior during our experiments. On the contrary, whenever the moths contacted the odor source they extended their proboscis, indicating feeding motivation. These conditions allowed us to test whether leaf odors play a role in guiding moths in their search for nectar. As expected, fewer moths contacted the odor sources emitting leaf odors compared to flower blends in the no-choice experiments. When we tested flower blends of D. wrightii and N. attenuata against a background of leaf volatiles emitted by conspecific plants, the moths preferred the combined odor over the flower blend alone. The behavioral activity of leaf odors is thus not restricted to oviposition, as leaf odors co-act with flower blends, thereby increasing the attractiveness of the flower blend. Raguso and Willis found in good agreement with our results that both vegetative and flower odors synergize visually guided feeding. Surrogate flowers were attractive and induced proboscis extension when scented with either flower or vegetative plant odors. However, in their experiments vegetative plant odors failed to enhance the attractiveness of scented surrogate flowers. A number of factors may explain this discrepancy. Raguso and Willis performed their experiments in the open field. Therefore, even the artificial flowers that were scented only with flower odor were presented against an olfactory background. Furthermore, the study was performed with wild moths that were most probably experienced. The moths’ preference for flower odors is flexible and influenced by learning. Wild moths may learn to associate the multimodal D. wrightii flower percept to a nectar reward. Furthermore, appearance, i.e. size, reflectance, and shape AbMole Acetylcorynoline determine the attractiveness of visual flower stimuli. While we used a small piece of filter paper as a minimalized visual stimulus these authors used true D. wrightii flowers or surrogate flowers of similar size.
show oviposition behavior and lay unfertilized eggs. However, in agreement with the data reported by we never observed abdomen curling, which would indicate egg-laying behavior during our experiments. On the contrary, whenever the moths contacted the odor source they extended their proboscis, indicating feeding motivation. These conditions allowed us to test whether leaf odors play a role in guiding moths in their search for nectar. As expected, fewer moths contacted the odor sources emitting leaf odors compared to flower blends in the no-choice experiments. When we tested flower blends of D. wrightii and N. attenuata against a background of leaf volatiles emitted by conspecific plants, the moths preferred the combined odor over the flower blend alone. The behavioral activity of leaf odors is thus not restricted to oviposition, as leaf odors co-act with flower blends, thereby increasing the attractiveness of the flower blend. Raguso and Willis found in good agreement with our results that both vegetative and flower odors synergize visually guided feeding. Surrogate flowers were attractive and induced proboscis extension when scented with either flower or vegetative plant odors. However, in their experiments vegetative plant odors failed to enhance the attractiveness of scented surrogate flowers. A number of factors may explain this discrepancy. Raguso and Willis performed their experiments in the open field. Therefore, even the artificial flowers that were scented only with flower odor were presented against an olfactory background. Furthermore, the study was performed with wild moths that were most probably experienced. The moths’ preference for flower odors is flexible and influenced by learning. Wild moths may learn to associate the multimodal D. wrightii flower percept to a nectar reward. Furthermore, appearance, i.e. size, reflectance, and shape AbMole Acetylcorynoline determine the attractiveness of visual flower stimuli. While we used a small piece of filter paper as a minimalized visual stimulus these authors used true D. wrightii flowers or surrogate flowers of similar size.