In plants, a subfamily of helix-hairpin-helix-Gly/Pro/Asp DNA glycosylases have been identified as demethylases involved in active cytosine DNA demethylation. These bifunctional glycosylases remove the 5-mC base and then cleave the DNA backbone at the resulting abasic site. Subsequent action by base excision repair machinery results in the replacement of 5mC with an unmethylated cytosine. Previous studies demonstrated the biological role of the glycosylase-mediated demethylation of DNA in Arabidopsis. Loss-of-function mutations in Demeter, a 5-mC DNA glycosylase gene in Arabidopsis, lead to impaired endosperm and embryo development, and eventually in seed abortion. Hypermethylation of cytosine occurred in genomes of plants that lack members of the DNA glycosylase demethylase family, e.g. repressor of silencing 1, DMElike 2, and DME-like 3. Mammals appear to lack the activity of glycosylases that can excise 5-mC specifically. However, active DNA demethylation may also be achieved in mammals through a BER pathway by DNA glycosylases, though it requires Coptisine-chloride oxidation of 5-mC as the first step. In this vein, 5-mC is converted into 5-hydroxymethylcytosine, 5-formylcytosine, and 5-carboxylcytosine by the ten-eleven translocation family of DNA dioxygenases through iterative oxidation. The mammalian TET proteins responsible for these oxidative modifications contain a catalytic domain that is typical of Fe and 2-oxoglutarate dependent dioxygenases, members of the cupin superfamily. By using computational Tubuloside-A analysis, Iyer et al. reported several distinct families of Fe and 2OG-dependent dioxygenases that are likely to be involved in oxidation of 5-mC in mammals and other early branching eukaryotes such as fungi and algae. In contrast, enzymes of this family have not been identified in multicellular plants. The prediction of lack of this cytosine modifying enzymatic activity in plants was strengthened by the fact that, despite some preliminary results reporting the presence of 5HmC, there is yet no definitive evidence supporting the presence of oxidation products of 5-mC in plants. As it was considered that this conclusion was based on the absence of evidence rather than the evidence of absence, we set out to assess the levels of these modified bases in Arabidopsis DNA using a liquid chromatography coupled with tandem mass spectrometry method. In this context, we recently reported the application of LC-MS/MS/MS, along with the isotope-dilution technique, for sensitive and accurate quantification of 5-hydroxymethyl-29-deoxycytidine in mammalian cells and tissues. Herein, we measured these four modified nucleosides along with the diastereomer of 8,59-cyclo-29-deoxyguanosine, a reliable biomarker for endogenously induced oxidative DNA damage, in Arabidopsis DNA. Our results demonstrated the presence of these modified nucleosides in Arabidopsis. Their relatively low 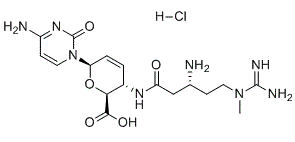 levels in genomic DNA, however, suggest that, in contrast to the observations made for mammals, they are not likely to be formed from enzyme-mediated oxidation reactions. The combination of LC-MS3 with the stable-isotope dilution technique provides a sensitive and accurate method for the measurement of oxidation products of 5-mdC, together with 5HmdU and S-cdG, in Arabidopsis tissues. To our knowledge, this is the first rigorous quantification of all three 5-mdC modification products in a flowering plant. Since the recent discoveries of Tetmediated oxidation of 5-mdC to 5-HmdC, 5-FodC, and 5-CadC, many biophysical and biochemical techniques have been employed for their detection, including LC-MS analysis, thin layer chromatography, chemical derivatization followed by sequencing analysis.
levels in genomic DNA, however, suggest that, in contrast to the observations made for mammals, they are not likely to be formed from enzyme-mediated oxidation reactions. The combination of LC-MS3 with the stable-isotope dilution technique provides a sensitive and accurate method for the measurement of oxidation products of 5-mdC, together with 5HmdU and S-cdG, in Arabidopsis tissues. To our knowledge, this is the first rigorous quantification of all three 5-mdC modification products in a flowering plant. Since the recent discoveries of Tetmediated oxidation of 5-mdC to 5-HmdC, 5-FodC, and 5-CadC, many biophysical and biochemical techniques have been employed for their detection, including LC-MS analysis, thin layer chromatography, chemical derivatization followed by sequencing analysis.